- Original article
- Open access
- Published:
600-ns pulsed electric fields affect inactivation and antibiotic susceptibilities of Escherichia coli and Lactobacillus acidophilus
AMB Express volume 10, Article number: 55 (2020)
Abstract
Cell suspensions of Escherichia coli and Lactobacillus acidophilus were exposed to 600-ns pulsed electric fields (nsPEFs) at varying amplitudes (Low-13.5, Mid-18.5 or High-23.5 kV cm−1) and pulse numbers (0 (sham), 1, 5, 10, 100 or 1000) at a 1 hertz (Hz) repetition rate. The induced temperature rise generated at these exposure parameters, hereafter termed thermal gradient, was measured and applied independently to cell suspensions in order to differentiate inactivation triggered by electric field (E-field) from heating. Treated cell suspensions were plated and cellular inactivation was quantified by colony counts after a 24-hour (h) incubation period. Additionally, cells from both exposure conditions were incubated with various antibiotic-soaked discs to determine if nsPEF exposure would induce changes in antibiotic susceptibility. Results indicate that, for both species, the total delivered energy (amplitude, pulse number and pulse duration) determined the magnitude of cell inactivation. Specifically, for 18.5 and 23.5 kV cm−1 exposures, L. acidophilus was more sensitive to the inactivation effects of nsPEF than E. coli, however, for the 13.5 kV cm−1 exposures E. coli was more sensitive, suggesting that L. acidophilus may need to meet an E-field threshold before significant inactivation can occur. Results also indicate that antibiotic susceptibility was enhanced by multiple nsPEF exposures, as observed by increased zones of growth inhibition. Moreover, for both species, a temperature increase of ≤ 20 °C (89% of exposures) was not sufficient to significantly alter cell inactivation, whereas none of the thermal equivalent exposures were sufficient to change antibiotic susceptibility categories.
Introduction
Bacterial contamination is recognized as a persistent and growing global concern for biotic and abiotic systems. To combat this threat, society has relied on three main bacterial inactivation methods: (1) Pharmaceutical (ex. antibiotics), (2) chemical (ex. antiseptics and disinfectants) and (3) physical (ex. heat, UV irradiation, cold plasma). Estimates from recent market research analyses indicate that these bacterial decontamination methods are so abundant, they globally comprise multibillion/million-dollar (US) industries at $46 billion (Watson 2019a), $5.5 billion (John 2017) and $149 million (Watson 2019b), respectively. Despite the large financial investments and modern technological advances bacteria continue to evolve and evade inactivation methods. Thus, novel bacterial control methods are urgently required.
One physical technique to inactivate microbes, which has been utilized for 100+ years (Sitzmann et al. 2017), is pulsed electric field(s) (PEF). Processes such as hospital effluent disinfection (Gusbeth et al. 2009), liquid and food pasteurization (Castro et al. 1993; Barbosa-Canovas et al. 2000), biofilm breakdown (Khan and El-Hag 2011; Freebairn et al. 2013) and topical burn antiseptic (Golberg et al. 2014) have found strong utilization for PEF. One of the main advantages for this inactivation process, versus the other available techniques, is the lack of resistance to the inactivation mode of action—membrane charging with subsequent, permanent breakdown of the cell wall (Katsuki et al. 2002; Dermol and Miklavčič 2017), also known as irreversible electroporation. However, the main disadvantage of this technology has been the large thermal gradient that is generated by the long (µs to > 1 s) pulses utilized, drastically restricting the use of this technology to heat-tolerant products. However, newly engineered nanosecond pulse generators, which create a significantly reduced thermal gradient, are now commercially available and have the potential to expand the PEF technology as a bacterial decontamination technique for heat-sensitive applications and products. This type of technology may prove especially useful for in vivo medical applications in which the subsequent heating of the surrounding tissue would be deleterious, as well as industrial food processing applications which require lower heat thresholds to maintain the sensory and physical properties of the food product (Barbosa-Canovas et al. 2000).
In this research study we examined the effects of a sub-microsecond pulse—600 ns, on two ubiquitous bacterial species, Escherichia coli and Lactobacillus acidophilus. We evaluated the impact of increasing the applied amplitude and pulse number to establish a range of inactivation thresholds at various exposure parameters. Given the effectiveness of nsPEF to inactivate bacteria, we hypothesized that the electroporated bacterial cells would be more sensitive to various antibiotics. To test this hypothesis, we exposed the treated cells to antibiotic-soaked discs and measured subsequent zones of growth inhibition. Furthermore, we measured the thermal gradient produced by each exposure and independently applied that thermal equivalent (TE) to both the inactivation and antibiotic susceptibility experiments. These results would validate that the observed inactivation and increase in antibiotic susceptibility was not based on thermal loading of the samples. We postulate that as a non-pharmaceutical, non-chemical, non-ionizing, and E-field driven technology, nsPEF has the potential to singularly or synergistically affect antibiotic treatment therapies, most especially antibiotic-resistant superbugs.
Materials and methods
Cell lines
Escherichia coli (ATCC 11775-MINI-PACK) and Lactobacillus acidophilus (ATCC 4357) cultures were purchased from American Type Culture Collection (ATCC) and propagated according to manufacturer’s protocol. Stocks were preserved by aliquoting respective bacterial broth with 50% glycerol (1:1) into 1.5 mL tubes and kept frozen at − 80 °C.
Exposure system
A Marx bank capacitor system (Fig. 1a), previously described (Ibey et al. 2014; Cantu et al. 2016), was used to generate the 600-ns unipolar pulse (Fig. 1b). A high-voltage probe (#P6015A Tektronix, Beaverton, OR, USA) connected to a high-speed oscilloscope (#TDS 30504B Tektronix, Beaverton, OR, USA) was used to measure the delivered pulse. For all exposures except 1000 pulses, the applied voltage from the high-voltage power supply was set at a fixed voltage to achieve the desired mean E-field amplitude in the cuvette (13.5 ± 0.6, 18.5 ± 2.5 and 23.5 ± 2.3 kV cm−1). For the 1000 pulse exposures, a rise in the temperature decreased the impedance of the cuvette resulting in a lower applied field. To maintain the mean exposure amplitude, the applied voltage was manually adjusted periodically. While this adjustment is not ideal, it ensured a more uniform E-field exposure, but likely contributed to some of the sample variability observed for 1000 pulse exposures. COMSOL Multiphysics® software v. 4.3b (COMSOL) was used to model E-field distribution within the exposure cuvette. The predicted E-field distribution displays uniformity throughout the exposure solution (Fig. 1c) thus the E-field is assumed to be spatially uniform throughout cellular exposures.

nsPEF exposure characterization. A schematic of the Marx bank capacitor system used to deliver the 600-ns PEF is provided (a). Representative oscilloscope traces of the applied voltages used to deliver an E-field amplitude of 13.5 (blue), 18.5 (black) or 23.5 (red) kV cm−1 (b) are shown. COMSOL Multiphysics® software was used to model the predicted E-field throughout the electroporation cuvette (both the x and y axis are equal in scale) (c)
Cell culture
Growth media was purchased from Becton, Dickson and Company (BD) (Franklin Lakes, NJ, USA). Nutrient broth (NB) (BD #213000) or De Man, Rogosa and Sharpe (MRS) broth (BD #288210) were utilized to culture E. coli and L. acidophilus, respectively. Overnight cultures were initiated by inoculating one vial from the − 80 °C stock into 50 mL of NB or MRS and incubated at 37 °C overnight. E. coli was cultured aerobically at 250 rpm. L. acidophilus was cultured in a capnophilic environment with elevated carbon dioxide levels using a BD GasPak 150 system (BD #260629), stationary. The following morning the optical density of a 100 µL aliquot was read in a spectrophotometer (#16032324 Synergy HTX BioTek, Winooski, VT, USA) set to a wavelength of 600 nm. To initiate experiments, a fresh 50 mL of growth media was inoculated with enough overnight culture to provide a starting optical density value of 0.05. Cultures were returned to their respective incubators and allowed to grow to log phase (approx. 4 h).
Exposures
To ensure consistency in exposure conditions both species were pelleted, washed and re-suspended in a 0.85% sodium chloride (NaCl) exposure solution (pH = 6.0, Conductivity = 1.33 S m−1). Final cell concentration was adjusted to approx. 1.5 × 108 cells mL−1. All cell solutions started at room temperature (approx. 22 °C).
- (i)
nsPEF induced temperature change (ΔT) (Thermal Gradient): A 400 µL sample of 0.85% NaCl solution was pipetted into a 2 mm aluminum, parallel plate electroporation cuvette (#89047-208 VWR International, Radnor, PA, USA). Prior to initiating the nsPEF exposure, a resting temperature (°C) of the NaCl solution was obtained by inserting a K-type thermocouple probe (#N1 USB-TC01 National Instruments, Austin, TX, USA) into the solution and then recorded using the N1 temperature logger software V1.1 (National Instruments, Austin, TX, USA). After the measurement, the probe was removed and the NaCl solution was exposed to a random series of: 1, 5, 10, 100 or 1000 pulses at a 1 Hz (one pulse/second) repetition rate and the amplitude was varied to 13.5, 18.5 or 23.5 kV cm−1. Sham exposures were performed by loading the cuvettes with NaCl solution and placing it into the pulser for 8 min (median time of the longest exposure) but no pulse was delivered. Immediately following exposure, the probe was re-inserted into the NaCl solution and the post exposure temperature was recorded. Change in temperature (ΔT) was obtained by subtracting the pre-exposure temperature from the post exposure temperature.
- (ii)
nsPEF: A 400 µL cell suspension (cells plus 0.85% NaCl) was pipetted into a 2 mm aluminum, parallel plate electroporation cuvette (#89047-208 VWR International, Radnor, PA, USA) and exposed to the above-mentioned exposure parameters.
- (iii)
nsPEF-induced thermal equivalent (TE) (heat stress): In total, three cell suspensions (cells suspended in 0.85% NaCl) of 120 µL each were pipetted into 3–200 µL PCR tubes (#TCS0803 BioRad, Hercules, CA, USA), capped (#TLS0801 BioRad, Hercules, CA, USA) and placed into an Eppendorf Mastercycler gradient thermocycler (#5331 Eppendorf, Hamburg, Germany). A resting temperature (°C) of the cell solution was obtained by inserting the thermocouple into a fourth PCR tube containing only 0.85% NaCl solution and recorded as mentioned above. The thermocycler was then set to the appropriate ΔT as observed in the thermal gradient experiment. Thermal exposure time was equivalent to nsPEF exposures (1 Hz) and was initiated once the ultimate temperature was reached in the thermocycler.
Enumeration of colony forming units (CFU)
Following the nsPEF or TE exposures, cells were removed from their respective exposure container and placed into a 1.5 mL tube. A tenfold serial dilution was performed by inoculating 900 µL of distilled water with a 100 µL aliquot from the previous dilution (no dilution to 106). Then, a 100 µL aliquot of the 104 to 106 samples were spread onto 20 mL of the respective growth agar using the spread plate technique. Following a 24 h incubation period, a direct colony count was taken. Log reduction of CFU was calculated by taking log10(Sham/Exposed). Lethal concentration (LC50) values were calculated using probit analysis in Excel; mean% dead values (across n experiments) were utilized to determine probit values.
Antibiotic susceptibility
A 100 µL aliquot of cell solution was removed from the no dilution Eppendorf tube (see above) and spread using the spread plate technique. The following antibiotic-soaked discs, purchased from BD, were subsequently stamped onto the inoculated E. coli and L. acidophilus plates: kanamycin (30 µg) (BD #B31301), tobramycin (10 µg) (BD #B31569), vancomycin (30 µg) (BD #B31353), ampicillin/sulbactam (10/10 µg) (BD #B31660), aztreonam (30 µg) (BD #B31641) and tetracycline (30 µg) (BD #B31344). The diameter of the zones of growth inhibition were measured (mm) following a 24 h incubation period and the bacteria were classified as:
Resistant (R): “Clinical efficacy has not been reliably shown in treatment studies” (Hardy Diagnostics 1996)
Intermediate (I): “Clinical applicability in body sites where the drug is physiologically concentrated or when a higher than normal dosage of the drug can be used. The MIC of the isolate may approach usually attainable blood and tissue levels but the response rate may be lower than for susceptible isolates” (Hardy Diagnostics 1996) or
Susceptible (S): “An infection due to the organism may be treated with the concentration of antimicrobial agent used, unless otherwise contraindicated” (Hardy Diagnostics 1996)
based on the Clinical and Laboratory Standards Institute (CLSI) pre-existing guidelines which were previously described by Hardy Diagnostics (1996) and Charteris et al. (1998).
Statistical analysis
Escherichia coli nsPEF experiments were performed 6 times (n = 6). All remaining experimental conditions (thermal gradient, nsPEF, TE, antibiotic susceptibility) were performed in three independent replicates (n = 3). Unless otherwise indicated, values represent the mean of n plus the standard error (SE). A student’s T-test with a p-value ≤ 0.05 was carried out for all experiments. A one-tail test was utilized for the thermal gradient experiments while a two-tail test was utilized for the nsPEF and TE experiments. A statistically significant difference is represented by an asterisk.
Results
Amplitude and pulse number affect 600-ns PEF thermal gradient
The temperature of the cell exposure solution (0.85% NaCl) was measured for each amplitude and pulse number used in this study (Fig. 2a–c). As expected, increasing the amplitude or pulse number resulted in an increase in the overall thermal gradient delivered to the sample. However, only two conditions: 1000 pulses, at 18.5 and 23.5 kV cm−1 produced a large enough ΔT capable of initiating irreversible bacterial inactivation (ΔT: 33 and 59 °C, respectively) (World Health Organization 2018; Bull et al. 2013).

nsPEF induced temperature change. Temperature change (ΔT) (°C) of 0.85% NaCl cell solution exposed to a 600-ns PEF at 13.5 (a), 18.5 (b) or 23.5 (c) kV cm−1 at 0 (sham), 1, 5, 10, 100 or 1000 pulses. Results are the mean ± SE of three independent experiments (n = 3). An asterisk represents a statistically significant difference from the sham
nsPEF inactivate E. coli and L. acidophilus to a greater extent than their thermal equivalents (TE)
The inactivation of E. coli and L. acidophilus, as measured by CFU, was evaluated after a 600-ns PEF or its TE at various amplitudes and pulse numbers.
- (i)
nsPEF exposures: For both E. coli (Fig. 3a–c, blue) and L. acidophilus (Fig. 4a–c, blue), inactivation was positively correlated to amplitude and/or pulse number. In general, cell inactivation increased as amplitude and/or pulse number increased. A maximum log reduction of CFU mL−1 was achieved at 1000 pulses for all amplitudes examined (a. 13.5 kV cm−1: 0.9, 0.5) (b. 18.5 kV cm−1: 2.5, 3.7) (c. 23.5 kV cm−1: 5.6, 6.9) for E. coli and L. acidophilus, respectively.
Fig. 3 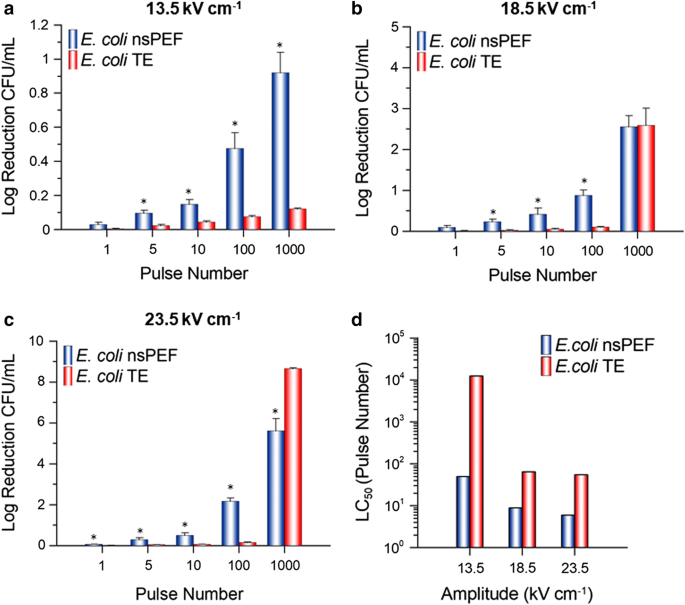
E. coli cell inactivation. Cell inactivation (log reduction CFU mL−1) (a–c) and LC50 values (d) for E. coli exposed to a 600-ns PEF (blue) or its respective TE (red) at an amplitude of 13.5, 18.5 or 23.5 kV cm−1 and 1, 5, 10, 100 or 1000 pulses. nsPEF results are normalized to the sham and are the mean plus SE of six independent experiments (n = 6). TE results are normalized to the sham and are the mean plus SE of three independent experiments (n = 3). LC50 values were calculated by utilizing the mean log reduction CFU mL−1 at each amplitude. An asterisk represents a statistically significant difference (p ≤ 0.05) between the nsPEF and TE exposure for that pulse number
Fig. 4 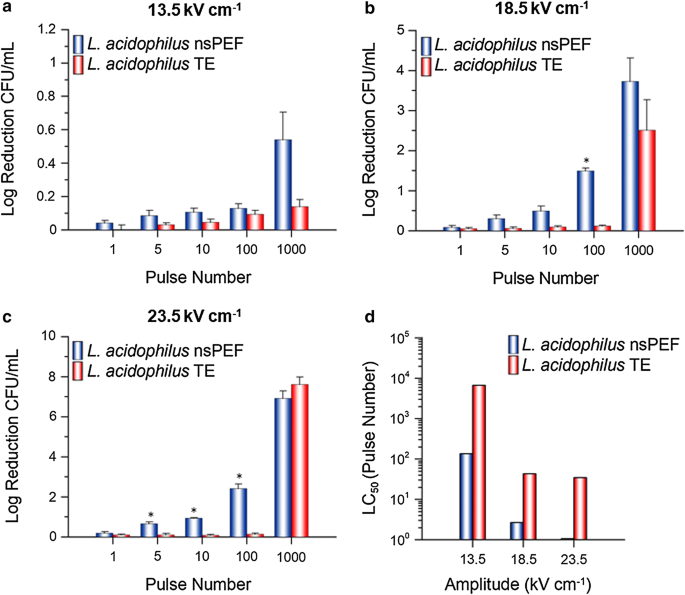
L. acidophilus cell inactivation. Cell inactivation (log reduction CFU mL−1) (a–c) and LC50 values (d) for L. acidophilus exposed to a 600-ns PEF (blue) or its respective TE (red) at an amplitude of 13.5, 18.5 or 23.5 kV cm−1 and 1, 5, 10, 100 or 1000 pulses. nsPEF and TE results are normalized to the sham and are the mean plus SE of three independent experiments (n = 3). LC50 values were calculated by utilizing the mean log reduction CFU mL−1 at each amplitude. An asterisk represents a statistically significant difference (p ≤ 0.05) between the nsPEF and TE exposure for that pulse number
- (ii)
TE exposures: To better understand if the observed increase in cell inactivation was due in-part to the nsPEF-induced thermal gradient, both species were subjected to a TE from each amplitude and pulse number examined (Figs. 3a–c and 4a–c). Only two exposure parameters: 1000 pulses at 18.5 and 23.5 kV cm−1 produced a substantial log reduction of CFU for E. coli (Fig. 3b, c, red) and L. acidophilus (Fig. 4b, c, red). However, this was expected as the thermal gradient experiment resulted in a change in temperature above the normal physiological range for both species. For the remainder of the exposures (88% of total exposures examined), log reduction CFU remained below 0.15, meaning ≥ 87% viability.


nsPEF increase the efficacy of various antibiotics on E. coli and L. acidophilus
The effectiveness of various antibiotics was examined for nsPEF and the TE exposures by measuring zones of inhibition following a 24 h incubation. Table 1 provides results for which the antibiotic susceptibility of the sham began as resistant (R) or intermediate (I), and the exposed sample transitioned to intermediate (I) or susceptible (S).
- (i)
E. coli: Zones of inhibition for kanamycin (30 µg) or tobramycin (10 µg) were larger when nsPEF was applied prior to the treatment. The kanamycin treated samples transitioned from intermediate to susceptible as early as 10 pulses in the 18.5 and 23.5 kV cm−1 exposures. “Susceptible” was also attained in the 13.5 kV cm−1 amplitude however, it was not achieved until 1000 pulses. The tobramycin treated samples transitioned from resistant to susceptible by 100 pulses in the 23.5 kV cm−1 exposure. Additionally, a transition from resistant to intermediate was observed in the 1000 pulse 18.5 kV cm−1 sample.
- (ii)
L. acidophilus: Zones of inhibition for vancomycin (30 µg) or ampicillin-sulbactam (10/10 µg) were larger when nsPEF was applied prior to the treatment. The vancomycin treated samples transitioned from resistant to intermediate as early as 5 pulses in the 23.5 kV cm−1 exposure. Intermediate susceptibility was also observed by the two lower amplitudes examined; however, it was not achieved until 1000 pulses. The ampicillin-sulbactam treated samples transitioned from intermediate to susceptible as early as 10 pulses in the 23.5 kV cm−1 exposure. “Susceptible” was also attained in the other two amplitudes examined however it was not achieved until 100 pulses.
For both species, a measurement for the 1000 pulse 23.5 kV cm−1 exposures could not be verified as there was no bacterial growth on the culture plate, indicative of total cell inactivation by either nsPEF, the thermal gradient or a combination of the two. Moreover, for all three amplitudes and four antibiotics reported in Table 1, none of the TE exposures were sufficient to change the susceptibility indicating that under these parameters’ inactivation was directly related to E-field effects and not a thermal gradient.
Discussion
In the present study we show in vitro results for the efficacy of 600-ns PEF to inactivate Escherichia coli and Lactobacillus acidophilus. While the effects of nsPEF have been studied on E. coli (Chalise et al. 2006; Perni et al. 2007; Guionet et al. 2014, 2015; Novickij et al. 2018) and other species such as Staphylococcus aureus (Chaturongakul and Kirawanich 2012; Vadlamani et al. 2018; Novickij et al. 2019), Salmonella typhimurium (Perni et al. 2007) and Bacillus subtilis (Katsuki et al. 2002); we chose to compare the effects of E. coli with L. acidophilus for two reasons. First, several studies have suggested that cell size and shape play a critical role in transmembrane potential charging and subsequent membrane pore formation (Kandušer and Miklavčič 2008; Khan and El-Hag 2011). Thus, to reduce effects resulting from differences in size and shape we chose two morphologically similar species; both species are rod shaped with similar size dimensions (Hardy Diagnostics 1996–2016; Reshes et al. 2008). Second, as we wanted to compare the effects of two physiologically different species (Gram negative and Gram positive), we chose the Gram-positive L. acidophilus, as it is generally considered to be apathogenic and moreover a probiotic. Furthermore, to our knowledge, we are the first group to report nsPEF inactivation results for L. acidophilus. Comparing the species examined here is important for potential applications where nsPEF exposures could be utilized to selectively target a bacterial species. For example, in vivo medical applications where a mixed population of pathogens and non-pathogens exist or in situ for food sterilization/liquid pasteurization purposes.
For comparison of cell inactivation thresholds between E. coli and L. acidophilus we utilized a range of exposure parameters including: E-field amplitude (Low-13.5, Mid-18.5 and High-23.5 kV cm−1) and pulse number (0 (sham), 1, 5, 10, 100 or 1000 pulses). This range of exposure conditions allowed us to determine parameter combinations that could be used to selectively target each species respectively. Furthermore, as studies have suggested that parameters of the exposure solution (ex. pH, conductivity) are equally important as cell type to electropermeabilization metrics (Kotnik et al. 1997; Stoodley et al. 1997), we chose to remove the cells from their respective growth media and re-suspend each cell type into a saline exposure solution; thus limiting potential differences based on differences in media composition. Our results show that, in general, L. acidophilus was more susceptible to the 600-ns PEF than E. coli; mean nsPEF LC50 values were higher for E. coli (Fig. 3d, blue) than L. acidophilus (Fig. 4d, blue) for both the 18.5 and 23.5 kV cm−1 exposures. However, this was not the case for the 13.5 kV cm−1 nsPEF exposures, suggesting that there may be an E-field threshold for inactivation of L. acidophilus.
These results were intriguing to us as many historical studies have suggested that based on differences in the electrical properties of the peptidoglycan layer for Gram-negative and Gram-positive species, a Gram-negative bacterium should be more susceptible to the effects of PEF (Hülsheger et al. 1983). However, a review of more recent PEF studies has proposed that this may not always be the case. For instance, García et al. (2005) examined the viability of various bacterial species to PEF treatments in an exposure medium with a pH of 4 or 7. Their results suggested that the same cell type will display inactivation differentially based on the pH of the exposure media (García et al. 2005). While the emphasis of this manuscript was on the effects of the E-field and temperature, we also examined the effect that the 600-ns PEF would have on the pH of the unbuffered NaCl exposure media. Utilizing the largest amplitude of 23.5 kV cm−1 we measured the final pH from all pulse numbers utilized, as this would display the most extreme pH changes. We observed the pH increase as the pulse number increased: 6.0 (sham), 6.3 (1 pulse), 6.5 (5 pulses), 6.8 (10 pulses), 7.9 (100 pulses) and 8.9 (1000 pulses). It is important to note that these pH changes are attributed to electrochemistry within the cuvette related to the aluminum electrodes and therefore would scale with amplitude. Therefore, the pH measurements included in this paper represent the worst-case scenario for the bacteria and would be significantly less for the other two lower amplitudes. However, for exposures at an amplitude of 23.5 kV cm−1 and pulses ≤ 10, the cumulative change in pH is rather modest, therefore the observed impact on cells would likely be independent of pH. For the 100 and 1000 pulse exposures, the pH change is significant. Although, it is important to note that this pH is achieved over the course of the pulse application and the bacteria only experience the extreme pH for a short duration between end of exposure and analysis. This is in contrast to García et al. (2005), where the bacteria are placed into a buffer already adjusted to a set pH level. Furthermore, for 1000 pulse exposures, where the pH shows its largest increase, temperature also increases making isolation of nsPEF-specific effects on the cells difficult. This is a topic that we would like to further investigate in future studies.
Furthermore, as it is theorized that inactivation is directly related to pore formation in the membrane (Tieleman et al. 2003; Thompson et al. 2016; Meglic and Kotnik 2016), our results correlate well to a study conducted by Piggot et al. (2011) in which molecular dynamic simulations of electroporation of the Gram-negative E. coli outer membrane and the Gram-positive S. aureus membrane were conducted (Piggot et al. 2011). In these simulations, the E. coli outer membrane was more resistant to poration than the S. aureus membrane; the higher resistance was attributed to reduced mobility of the lipopolysaccharide molecules such that the phospholipids were required to fill the water-filled pore resulting in lipid flip-flop (Piggot et al. 2011). The results from that simulation study suggest that other Gram-positive species, such as L. acidophilus, may be susceptible to electroporation and subsequent inactivation more easily than E. coli. Nevertheless, our results indicate that under these exposure conditions it was ultimately the total energy delivered (amplitude, pulse number and pulse duration) that determined the extent of cell inactivation for an individual species.
Although nsPEF technology is largely considered to be E-field driven, during treatment there is an energy-dependent increase in process temperature due to electric current flow and individual product resistance (Schottroff et al. 2018); thus, additional inactivation effects due to heating should be considered. Consequently, for temperature-sensitive applications, it was important to differentiate inactivation triggered by the E-field from the thermal gradient. Additionally, beneficial compounds (e.g. antimicrobial agents, nutrients, etc.) found within target materials and their physical properties may also be affected by temperature gradients induced by extreme PEF exposures. Although the greatest extent of cell inactivation was achieved by 1000 pulses at 18.5 and 23.5 kV cm−1; for temperature-sensitive applications, we postulate that the more suitable exposures would be ≤ 100 pulses. At these pulse numbers a significant amount of cell inactivation was obtained by the nsPEF exposure while the thermal gradient and subsequent TE inactivation was negligible. Furthermore, the 13.5 kV cm−1 amplitude produced a greater inactivation difference between E. coli and L. acidophilus, while greatly restricting the thermal gradient, at 100 and 1000 pulses. These parameters might be especially useful to target one cell type over the other. However, an important item to note from this study is that the results from the TE exposures reflect a more severe result than what the nsPEF exposed cells actually experienced. This difference can mainly be attributed to the manner by which the nsPEF or TE cells were delivered heat. For instance, during the nsPEF exposures a 1 Hz repetition rate was directly correlated to exposure time (i.e. it took 5 s to deliver 5 pulses). However, for the TE exposures, there was a rate differential by which the thermocycler would adjust temperature gradients between “high” and “low” temperatures. For example, exposures which were set to a higher temperature (ex. 100 or 1000 pulses) were heated more quickly than samples that were set to a lower temperature (ex. 1, 5, or 10 pulses). Therefore, to maintain consistency throughout the experiment, the exposure time (equivalent to 1 Hz) was only initiated after a final temperature was achieved. This resulted in a longer total exposure time for the TE exposures as compared to the nsPEF exposures. Interestingly, although the TE LC50 values show slight variation at the different amplitudes between E. coli (Fig. 3d, red) and L. acidophilus (Fig. 4d, red), a similar trend can be observed in which the low amplitude exposures (13.5 kV cm−1) require more pulses to kill 50% of the population. However, once a heat threshold was met by pulse number the applied amplitude (18.5 vs. 23.5 kV cm−1) became irrelevant.
Lastly, as there is an urgent need for novel techniques to combat antibiotic resistant microbes (CDC 2019), we investigated the possible synergistic effects that nsPEF or the TE exposures could have on antibiotic susceptibilities. To examine this, we exposed both the nsPEF and TE exposed cells to four classes (aminoglycoside, tetracycline, glycopeptide and β-lactams) of antibiotic soaked disks and allowed the bacteria to grow overnight as the antibiotic diffused throughout the media. Results in Table 1 show that multiple nsPEF exposures can enhance the susceptibility of antibiotics for both E. coli and L. acidophilus, indicative by increased zones of growth inhibition. We also display results for antibiotics that, for E. coli, originated as susceptible but displayed increased zones of inhibition for select nsPEF exposures (Additional file 1: Table S1). Although there were some minor increases in the zones of inhibition for the TE exposures, none of these treatments were significant enough to provide a change in susceptibility, indicating that the change in antibiotic susceptibility was due to the effects of the E-field and not to any induced heating. Although the different classes of antibiotics work on various cellular targets to inactivate bacteria (Li et al. 2015; Kapoor et al. 2017), the first step in the mechanism of action for all antibiotics is to traverse the cell membrane and enter the cytoplasm (James et al. 2009; Krause et al. 2016). Research conducted by Pillet et al. (2016) showed scanning electron, transmission electron and atomic force microscopy results demonstrating the morphological, mechanical and physical damage to the cell wall of Bacillus pumilus from various strengths of µs electric pulses (Pillet et al. 2016). These results strongly support the theory that PEF directly impact bacterial cell wall integrity. Therefore, we would conclude that damage to the integrity of the cell wall and nsPEF-induced poration of the membrane is reducing or removing this first step allowing for the translocation of the antibiotic to be easier, faster and thus more effective. However, these results were limited to testing the bacterial susceptibility to various antibiotics only after the nsPEF exposure was delivered. It is likely that the addition of antibiotics to the pulsing medium could have increased the efficiency of cell death, but it would have also introduced the possibility of direct electrotransfer of the antibiotic into the cell, which was intentionally avoided in this study. Future research should evaluate if electrotransfer of antibiotics during nsPEF exposure increases the level of death across the various bacterial species.
In conclusion, this study has shown the efficacy of 600-ns PEF, at various amplitudes and pulse numbers, to inactivate cells and enhance the susceptibility of E. coli and L. acidophilus to various antibiotics. These results have aided in further building a foundation for the utilization of nsPEF in heat-sensitive bacterial inactivation applications and as a possible tool to combat antibiotic resistant microbes. Based on the positive results of this study, future investigations into the utilization of nsPEF for decontamination with medically relevant anaerobic species like Clostridium difficile or species with special pathogenic traits like the capsular polysaccharide matrix of Klebsiella should also be investigated. Finally, although there does not currently seem to be resistance mechanisms by the bacteria to evade PEF inactivation, there is limited research on the topic. Future studies should include an exhaustive attempt to further understand bacterial resistance to PEF.
Availability of data and materials
Corresponding author could provide all the experimental data on valid request.
Abbreviations
- ns:
-
Nanosecond
- nsPEF:
-
Nanosecond pulsed electric field(s)
- kV:
-
Kilovolts
- cm:
-
Centimeter
- Hz:
-
Hertz
- E-field:
-
Electric field
- h:
-
Hour
- PEF:
-
Pulsed electric field(s)
- µs:
-
Microsecond
- s:
-
Second
- TE:
-
Thermal equivalent
- mL:
-
Milliliter
- C:
-
Centigrade
- BD:
-
Becton, Dickson and Company
- NB:
-
Nutrient broth
- MRS:
-
De Man, Rogosa and Sharpe
- rpm:
-
Rotations per minute
- µL:
-
Microliter
- nm:
-
Nanometer(s)
- approx:
-
Approximately
- NaCl:
-
Sodium chloride
- pH:
-
Potential hydrogen
- S/m:
-
Siemens per meter
- Δ:
-
Delta or change in
- mm:
-
Millimeter
- PCR:
-
Polymerase chain reaction
- CFU:
-
Colony forming unit(s)
- LC50 :
-
Lethal concentration 50%
- µg:
-
Micrograms
- R:
-
Resistant
- I:
-
Intermediate
- S:
-
Susceptible
- SE:
-
Standard error
References
Barbosa-Canovas GV, Pierson MD, Zhang QH, Schaffner DW (2000) Special supplement kinetics of microbial inactivation for alternative food processing technologies (pulsed electric fields). J Food Sci 65:65–79
Bull M, Plummer S, Marchesi J, Mahenthiralingam E (2013) The life history of Lactobacillus acidophilus as a probiotic: a tale of revisionary taxonomy, misidentification and commercial success. FEMS Microbiol Lett 349:77–87
Cantu JC, Tarango M, Beier HT, Ibey BL (2016) The biological response of cells to nanosecond pulsed electric fields is dependent on plasma membrane cholesterol. Biochim Biophys Acta Biomembr 1858:2636–2646
Castro AJ, Barbosa-Canovas GV, Swanson BG (1993) Microbial inactivation of foods by pulsed electric fields. J Food Process Preserv 17:47–73
CDC (2019) Antibiotic resistance threats in the United States. CDC. https://www.cdc.gov/drugresistance/pdf/threats-report/2019-ar-threats-report-508. Accessed 21 Oct 2019
Chalise PR, Perni S, Shama G, Novac BM, Smith IR, Kong MG (2006) Lethality mechanisms in Escherichia coli induced by intense sub-microsecond electrical pulses. Appl Phys Lett 89:153902
Charteris WP, Kelly PM, Morelli L, Collins JK (1998) Antibiotic susceptibility of potentially probiotic Lactobacillus species. J Food Prot 61:1636–1643
Chaturongakul S, Kirawanich P (2012) An in vitro study of bacterial survival in response to high-intensity nanosecond pulse stimulation. In: 2012 9th international conference on electrical engineering/electronics, computer, telecommunications and information technology, ECTI-CON 2012
COMSOL Multiphysics® v. 4.3b. http://www.comsol.com. COMSOL AB, Stockholm, Sweden
Dermol J, Miklavčič D (2017) Mathematical models describing cell death due to electroporation. In: Miklavčič D (ed) Handbook of electroporation. Springer, Cham, pp 1199–1218
Freebairn D, Linton D, Harkin-Jones E, Jones DS, Gilmore BF, Gorman SP (2013) Electrical methods of controlling bacterial adhesion and biofilm on device surfaces. Expert Rev Med Devices 10:85–103
García D, Gómez N, Raso J, Pagán R (2005) Bacterial resistance after pulsed electric fields depending on the treatment medium pH. Innov Food Sci Emerg Technol 6:388–395
Golberg A, Broelsch GF, Vecchio D, Khan S, Hamblin MR, Austen WG, Sheridan RL, Yarmush ML (2014) Eradication of multidrug-resistant A. baumannii in burn wounds by antiseptic pulsed electric field. Technology 02:153–160
Guionet A, Joubert-Durigneux V, Packan D, Cheype C, Garnier JP, David F, Zaepffel C, Leroux RM, Teissié J, Blanckaert V (2014) Effect of nanosecond pulsed electric field on Escherichia coli in water: inactivation and impact on protein changes. J Appl Microbiol 117:721–728
Guionet A, David F, Zaepffel C, Coustets M, Helmi K, Cheype C, Packan D, Garnier JP, Blanckaert V, Teissié J (2015) E. coli electroeradication on a closed loop circuit by using milli-, micro- and nanosecond pulsed electric fields: comparison between energy costs. Bioelectrochemistry 103:65–73
Gusbeth C, Frey W, Volkmann H, Schwartz T, Bluhm H (2009) Pulsed electric field treatment for bacteria reduction and its impact on hospital wastewater. Chemosphere 75:228–233
Hardy Diagnostics (1996) HardyDisk™ antimicrobial sensitivity test instructions for use. https://catalog.hardydiagnostics.com/cp_prod/Content/hugo/HardyDiskASTProceduresandChart.pdf. Accessed 20 June 2018
Hardy Diagnostics (1996–2016) Lactobacillus. Catalog.hardydiagnostics. https://catalog.hardydiagnostics.com/cp_prod/Content/hugo/Lactobacillus.htm. Accessed 5 Nov 2019
Hülsheger H, Potel J, Niemann EG (1983) Electric field effects on bacteria and yeast cells. Radiat Environ Biophys 22:149–162
Ibey BL, Ullery JC, Pakhomova ON, Roth CC, Semenov I, Beier HT, Tarango M, Xiao S, Schoenbach KH, Pakhomov AG (2014) Bipolar nanosecond electric pulses are less efficient at electropermeabilization and killing cells than monopolar pulses. Biochem Biophys Res Commun 443:568–573
James CE, Mahendran KR, Molitor A, Bolla JM, Bessonov AN, Winterhalter M, Pagès JM (2009) How β-lactam antibiotics enter bacteria: a dialogue with the porins. PLoS ONE 4:e5453
John J (2017) global antiseptics and disinfectants market will reach USD 8.10 Billion by 2021: Zion Market Research. Globenewswire. https://www.globenewswire.com/news-release/2017/05/12/984235/0/en/Global-Antiseptics-and-Disinfectants-Market-will-reach-USD-8-10-Billion-by-2021-Zion-Market-Research.html. Accessed 13 Nov 2019
Kandušer M, Miklavčič D (2008) Electroporation in biological cell and tissue: an overview. In: Food engineering series, pp 1–37
Kapoor G, Saigal S, Elongavan A (2017) Action and resistance mechanisms of antibiotics: a guide for clinicians. J Anaesthesiol Clin Pharmacol 33:300
Katsuki S, Moreira K, Dobbs F, Joshi RP, Schoenbach KH (2002) Bacterial decontamination with nanosecond pulsed electric fields. In: IEEE conference record of power modulator symposium
Khan OGM, El-Hag AH (2011) Biological cell electroporation using nanosecond electrical pulses. J Med Imaging Health Inform 1:278–283
Kotnik T, Bobanović F, Miklavčič D (1997) Sensitivity of transmembrane voltage induced by applied electric fields—a theoretical analysis. Bioelectrochem Bioenerg 43:285–291
Krause KM, Serio AW, Kane TR, Connolly LE (2016) Aminoglycosides: an overview. Cold Spring Harb Perspect Med 6:a027029
Li F, Collins JG, Keene FR (2015) Ruthenium complexes as antimicrobial agents. Chem Soc Rev 44:2529–2542
Meglic SH, Kotnik T (2016) Electroporation-based applications in biotechnology. In: Miklavcic D (ed) Handbook of electroporation. Springer, Cham, pp 1–17
Novickij V, Zinkevičienė A, Stanevičienė R, Gruškienė R, Servienė E, Vepštaitė-Monstavičė I, Krivorotova T, Lastauskienė E, Sereikaitė J, Girkontaitė I, Novickij J (2018) Inactivation of Escherichia coli using nanosecond electric fields and nisin nanoparticles: a kinetics study. Front Microbiol 9:3006
Novickij V, Lastauskiene E, Staigvila G, Girkontaite I, Zinkevičiene A, Švediene J, Paškevičius A, Markovskaja S, Novickij J (2019) Low concentrations of acetic and formic acids enhance the inactivation of Staphylococcus aureus and Pseudomonas aeruginosa with pulsed electric fields. BMC Microbiol 19:73
Perni S, Chalise PR, Shama G, Kong MG (2007) Bacterial cells exposed to nanosecond pulsed electric fields show lethal and sublethal effects. Int J Food Microbiol 120:311–314
Piggot TJ, Holdbrook DA, Khalid S (2011) Electroporation of the E. coli and S. aureus membranes: molecular dynamics simulations of complex bacterial membranes. J Phys Chem B 115:13381–13388
Pillet F, Formosa-Dague C, Baaziz H, Dague E, Rols MP (2016) Cell wall as a target for bacteria inactivation by pulsed electric fields. Sci Rep 6:19778
Reshes G, Vanounou S, Fishov I, Feingold M (2008) Cell shape dynamics in Escherichia coli. Biophys J 94:251–264
Schottroff F, Fröhling A, Zunabovic-Pichler M, Krottenthaler A, Schlüter O, Jäger H (2018) Sublethal injury and Viable but Non-culturable (VBNC) state in microorganisms during preservation of food and biological materials by non-thermal processes. Front Microbiol 9:2773
Sitzmann W, Vorobiev E, Lebovka N (2017) Pulsed electric fields for food industry: historical overview. In: Miklavčič D (ed) Handbook of electroporation. Springer, Cham, pp 2335–2354
Stoodley P, DeBeer D, Lappin-Scott HM (1997) Influence of electric fields and pH on biofilm structure as related to the bioelectric effect. Antimicrob Agents Chemother 41:1876–1879
Thompson GL, Roth CC, Kuipers MA, Tolstykh GP, Beier HT, Ibey BL (2016) Permeabilization of the nuclear envelope following nanosecond pulsed electric field exposure. Biochem Biophys Res Commun 470:35–40
Tieleman DP, Leontiadou H, Mark AE, Marrink SJ (2003) Simulation of pore formation in lipid bilayers by mechanical stress and electric fields. J Am Chem Soc 125:6382–6383
Vadlamani A, Detwiler DA, Dhanabal A, Garner AL (2018) Synergistic bacterial inactivation by combining antibiotics with nanosecond electric pulses. Appl Microbiol Biotechnol 102:7589–7596
Watson J (2019a) Global antibiotics market to reach USD 63.34 billion by 2026 | Reports and data. Globenewswire. https://www.globenewswire.com/news-release/2019/05/06/1817677/0/en/Global-Antibiotics-Market-To-Reach-USD-63-34-Billion-By-2026-Reports-And-Data.html. Accessed 13 Nov 2019
Watson J (2019b) Food and beverage disinfection market to reach USD 206.4 million by 2026 | reports and data. Globenewswire. https://www.globenewswire.com/news-release/2019/09/25/1920849/0/en/Food-and-Beverage-Disinfection-Market-To-Reach-USD-206-4-Million-By-2026-Reports-And-Data.html. Accessed 13 Nov 2019
World Health Organization (2018) E. coli. WHO. https://www.who.int/news-room/fact-sheets/detail/e-coli. Accessed 09 Oct 2019
Acknowledgements
The authors would like to thank the following member of the United States Air Force Research Laboratory Radio Frequency Bioeffects Branch for their assistance in laboratory set-up, data collection and/or manuscript preparation: Ms. Kaitlin Nelson, Mr. Zach Coker, Mr. Jason Payne, TSgt Nicholas Berardino and TSgt Timothy Muse.
Funding
This research was funded by the Air Force Office of Scientific Research Grant No. 16RHCOR348
Author information
Authors and Affiliations
Contributions
SM, RB, CR, and BI conceived and designed research. SM, SK, PT and RB conducted experiments. SM wrote the manuscript. All authors edited the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The author declares that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Additional file 1: Table S1.
Zones of growth inhibition for E. coli.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Martens, S.L., Klein, S., Barnes, R.A. et al. 600-ns pulsed electric fields affect inactivation and antibiotic susceptibilities of Escherichia coli and Lactobacillus acidophilus. AMB Expr 10, 55 (2020). https://doi.org/10.1186/s13568-020-00991-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13568-020-00991-y